|
Laboratory of Dr Enrique López-Juez |
Biological antennae also need to be tuned...
How light and plastid signals regulate leaf and chloroplast development
|
|
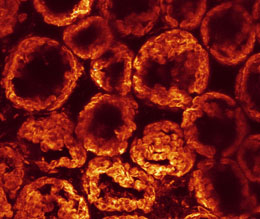
An optical section of a live Arabidopsis leaf, viewed from the top, under fluorescence excitation with blue light. Each leaf palisade cell (in this case very elongated because of growth at high light) appears as a tunnel coated internally by red-fluorescing chloroplasts. |
There is something fascinating (as many textbooks will tell us) about the study of plants and their response to the environment: since plants are sessile, they have to sense and adapt to their environment (a given individual cannot walk away from it!). And since light, through photosynthesis, is to plants what food is to animals, it is not surprising that exquisite mechanisms have evolved for plants to monitor their light environment. The study of those mechanisms is broadly known as "photomorphogenesis". Perhaps more surprising is the limited attention that the role those photomorphogenic mechanisms play in the development and performance of the photosynthetic machinery (leaf cells and their chloroplasts) has attracted. This is where our lab would like to make a contribution: the mechanisms through which photoreceptors control the development of chloroplasts and leaves, or in other words, the interface between "photomorphogenesis" and "photosynthesis".
We primarily use a combined physiological and genetic approach, utilising the most widely used plant model species, Arabidopsis thaliana.
Here are three areas we are trying to address. Click on any of them to jump to the relevant section below.
Links to photobiology and Arabidopsis
|
|
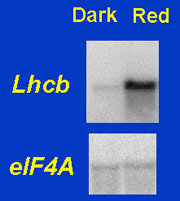
Chloroplasts do not develop in the dark or when their precursor plastids are damaged. The control of chloroplast biosynthesis is effected primarily at the level of expression of nuclear (and plastidic) genes which encode chloroplast proteins. One such group of genes is the Lhcb1 class, which encodes a major component of the photosynthetic light-harvesting complex. We are studying how phytochrome activates the expression of Lhcb1. |

|
We are trying in particular to assess whether the control by light of nuclear photosynthetic gene expression can ultimately be explained by the same control exerted by the functionality of plastids (the so called "plastid signal"). We have previously obtained evidence suggesting that this may be the case. |
|
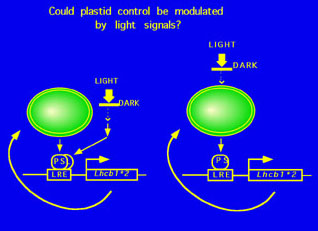
|
|
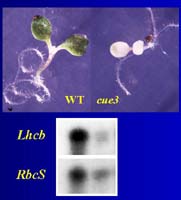
In the past, in the laboratory of J. Chory, we isolated Cab (Lhcb)-underexpresser (cue) mutants following the strategy described by Li et al. (1995), Plant Cell 7, 1599-1610. |

|
Several of these mutants show a correlation between the degree of plastid development (as shown for cue6) and the expression of these genes for photosynthetic components. Ultimately we are using the mutants as tools to answer the question: Do plastids play a direct role in transducing the light signals regulating their development? |
|
We are further trying to identify new mutants specifically in the control by phytochrome of Lhcb expression, using a reporter gene, green fluorescent protein (GFP) expressed under Lhcb control.
|
|
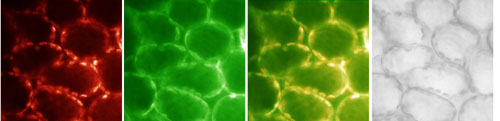
|
While trying to understand the role of plastid-to-nuclear signals in phytochrome induction of Lhcb, we generated mutants between hy1, a phytochrome chromophore mutant, and gun (genome uncoupled) mutants, in which plastid signals appear constitutively active, so that Lhcb expression is not shut down when plastids are dysfunctional. To our surprise, we found a strong interaction between hy1 and some, but not all, gun mutants, suggesting that at least one plastid signal was related to a group of plastid metabolites, tetrapyrroles (both chlorophyll and the phytochrome chromophore are tetrapyrroles). |
|
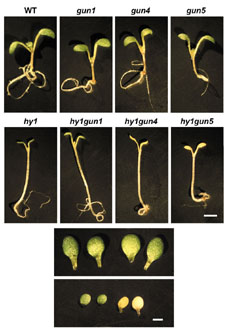
|
|
We are also addressing directly the relation between photosynthetic gene expression and the presence/absence of plastids, in other words, the basic positive or negative nature of the plastid signal. If photosynthetic genes are on when plastids are functional and off when they are dysfunctional, how would they be in the total absence of plastids? For the first time we can monitor this, since single cells devoid of plastids occur in the arc6 mutant (Pyke et al. 1994, Plant Physiol. 106, 1169-1177). This mutant is defective in plastid division and about one in four guard cells appears to be plastid-less. GFP, a fluorescent reporter gene product, allows us to monitor the expression of photosynthetic genes in those cells. This work is a collaboration with the laboratories of Kevin Pyke (University of Nottingham) and John Gray (University of Cambridge). We have already shown that photosynthetic genes are expressed (as monitored by GFP) specifically in photosynthetic cells, both mesophyll and guard cells. |
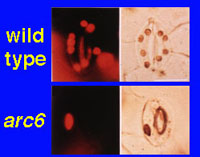
|
|
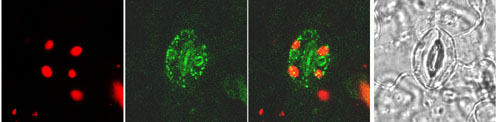
|
3. Light quantity and the control of leaf and chloroplast development |
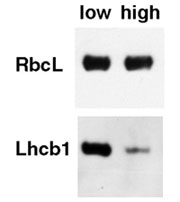
One critical challenge in the environment for leaves and their chloroplasts is the quantity of light they experience (because photosynthesis has to harvest light efficiently, but light can also become too much of a good thing). The fine-tuning (although it can become dramatic) of the photosynthetic machinery to the prevailing light level is known as "photosynthetic acclimation". It has become apparent that some acclimatory responses are mediated by light perception and processing by the photosynthetic apparatus itself. Those responses can also be triggered by changes in factors other than light. How this is brought about is, however, unclear. In addition, our evidence indicates that other responses are particularly sensitive to the irradiance of light, particularly of blue light. This suggests the involvement of a specific blue light photoreceptor. We are among a few labs who are trying to develop Arabidopsis, with its fully available genome sequence, as an organism with which to fully understand the mechanisms of acclimation.
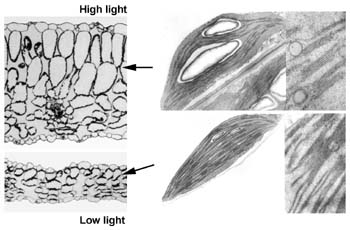
Among the things we know so far is that, in spite of the blue-dependence of the modulation of RbcS expression, a component of acclimation, all known blue photoreceptor single mutants in Arabidopsis (mutants in two cryptochromes and phototropin), as well as a double cryptochrome mutant, acclimate normally. We have recently constructed a triple blue photoreceptor mutant. Should this triple mutant acclimate normally, this would be evidence for the existence a photoreceptor yet to be uncovered.
The real protagonists (an incomplete list) to whom the credit belongs.

Chloroplasts are the primary factories of most of the global biomass. There is a basic intellectual need for understanding their biogenesis. Besides, there are vast possible applications of knowledge gained about them. Since chloroplast developmental regulation evolved under conditions of natural competition, not under monoculture as happens today in crops, there is potential for rational improvements in the performance of crops. Furthermore, chloroplasts are the most active biosynthetic sites in plants, and altering chloroplast biogenesis would alter the biosynthetic capacity of the plant overall.
Dreams to strive towards keep (us, at least) from stalling.
- E.López-Juez, contact details and short summary of the above research
- K. Pyke's lab, plastid developmental biology
- J. Chory's lab, light, hormones and plant development
- J. Gray's lab, chloroplast biogenesis:
- J. Haseloff's lab, GFP in Arabidopsis
- A. Millar's lab, light, rhythms and Lhcb
- E. Tobin's lab, light, plants and genes
- Peter Horton's lab, regulation of photosynthesis in the environment
- Nottingham Arabidopsis Stock Centre
- TAIR, everything Arabidopsis
- BL2020, teaching materials
![]()
©RHUL - 31 Jan 2001 - GJW